Fungal pathogen isolates and host plants
Maintenance of M. oryzae and its growth medium followed previously established protocols. The wild-type strain Guy11 was grown on complete medium in a growth chamber set to 25°C, with a 12-hour light–dark cycle for 10 days prior to collecting conidia for rice plant inoculation. Mycelial growth of various M. oryzae strains was assessed under different stress conditions on complete agar medium, with or without additional chemicals for a duration of 10 days. The chemicals used included 5 mM H2O2 for oxidative stress, 200 μg/ml congo red or calcofluor white for cell wall stress, and 0.01% SDS to study cell membrane stress.
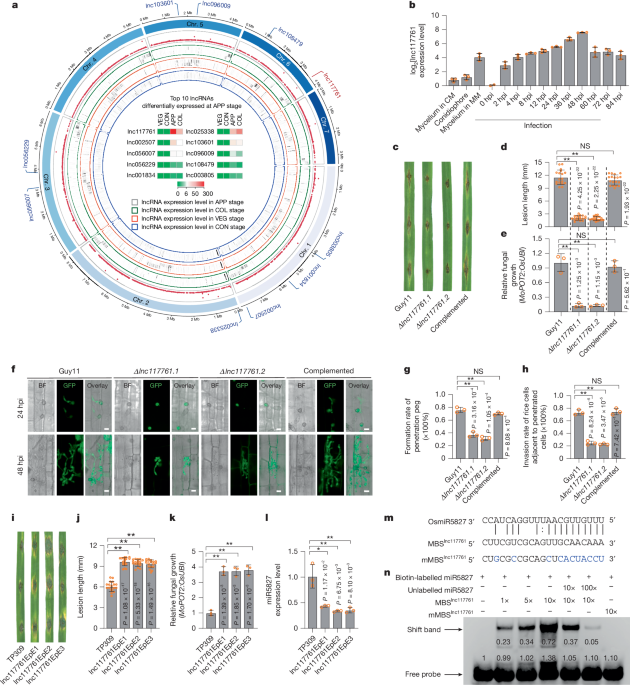
Infection of rice plants with M. oryzae was performed as described in prior studies. Conidial suspensions (1 × 105 spores/ml) were applied to rice plants, which were then incubated for 5 to 7 days before photographs were taken. The relative amount of fungal DNA was determined using the threshold cycle value (CT) of M. oryzae POT2 DNA compared to that of rice ubiquitin genomic DNA. The infection-related development of M. oryzae was analyzed based on earlier research at specified hours post-inoculation (hpi). The frequency of plant penetration was evaluated by counting the formation of appressoria or penetration pegs on rice leaf sheaths from the TP309 japonica cultivar or its transgenic lines, examining at least 50 spores in triplicate with an Axio Imager A2 microscope (Zeiss). hpi indicates hours post-inoculation with M. oryzae.
The R. solani AG1 strain was grown on potato dextrose agar plates at 28°C before mycelia were collected for inoculation following established reports. The F. graminearum PH-1 strain was routinely cultured on potato dextrose agar at 25°C, and mutants were created for infection assays on wheat seedlings as described in previous work. In this setup, TP309 rice was inoculated with M. oryzae and R. solani, while the wheat cultivar Nanmai 660 was inoculated with F. graminearum.
DNA and RNA manipulation
DNA extraction from fungi or plant samples was performed using the cetyltrimethylammonium bromide method, as previously detailed. RNA isolation followed established protocols where samples were ground into powder using liquid nitrogen and then transferred to RNase-free tubes. TRIzol reagent was employed for total RNA extraction as per manufacturer’s directions. RNA from M. oryzae was obtained from various sources: vegetative mycelia grown in liquid medium, conidiophores, appressoria, and invasive hyphae colonizing rice leaves. Reverse transcription was carried out with the total RNA using the HiScript III RT SuperMix for qPCR kit, followed by amplification on a NextGene QL96 instrument with standardized procedures.
RNA-seq analysis of M. oryzae
The wild-type strain Guy11 of M. oryzae was cultivated under four distinct conditions before total RNA extraction using an RNeasy Plant Mini Kit. Samples were collected from vegetative mycelia, conidia, appressoria, and invasive hyphae, with two biological replicates for each condition. RNA-seq library construction and sequencing analysis were carried out based on established methods. Genes that showed significant differential expression were identified by adjusted P values.
Generation of M. oryzae mutants
A split-marker deletion method via PCR was utilized to target the gene lnc117761 for deletion. Specifically, a 600-bp DNA segment of lnc117761 was replaced with a selection marker gene HPT, which provides resistance to hygromycin B. For genetic complementation, the full-length lnc117761 gene expression vector containing its native promoter was constructed. Additional variants were also developed to analyze the gene’s non-coding properties. Several lnc117761 constructs carrying nucleotide mutations were introduced into Δlnc117761 for functional analysis.
Generation of transgenic rice lines
Transgenic rice lines were created using a CRISPR–Cas9 method to disrupt miR5827. Two oligonucleotides targeting the OsmiR5827 precursor were selected for this purpose. Following customary procedures, genomic DNA samples from the transgenic lines were isolated and analyzed for targeted deletions. Rice lines with disrupted PKR1 were also created similarly, with specific mutations designed to eliminate the miR5827-targeting site. Overexpression of M. oryzae lnc117761 in rice utilized the CaMV 35S promoter in a binary vector to control expression.
Analysis of plant miRNA and gene expression levels
For miRNA level analysis, total RNA was extracted from various plant tissues including barley, A. thaliana, potato, B. distachyon, and wheat. Following standard extracting protocols, RNA was reverse transcribed with specific primers. For gene expression, RT–qPCR was carried out using specific kits, with the UBIQUITIN gene acting as a reference. Potential target genes of miR5827 were tabulated, followed by confirmation through RT–qPCR, ensuring specificity through proper controls. Each assay was repeated for reliability.
In situ hybridization assay
In situ hybridization was performed to determine the distribution pattern of lnc117761. Leaf sheaths from the TP309 rice cultivar were inoculated with M. oryzae and processed for microscopic analysis post-infection. The sections were fixed, embedded, and processed following standard protocols for in situ hybridization, utilizing specific probes to detect lnc117761 and control RNAs separately.
Detection of lnc117761 secretion
Multiple approaches were used to investigate the transport of lnc117761 from M. oryzae to rice. In one method, rice leaves around the infection site were sampled for analysis. In another, vesicles from M. oryzae were prepared and analyzed for lnc117761 distribution. A third approach involved transforming a GFP-expressing strain with an lnc117761–4×Pepper fusion construct for visualization.
Analyses of extracellular vesicles
Isolated extracellular vesicles from M. oryzae were examined using various methods, including transmission electron microscopy and nanoparticle tracking analysis. A series of steps were followed for sample preparation to ensure the integrity and appropriate labeling of the vesicles for imaging and analysis.
GFP reporter assay for miR5827 binding with lnc117761 or PKR1
A GFP reporter assay was used to evaluate the interaction of miR5827 with both lnc117761 and PKR1. Various constructs were prepared and transfected into plant tissues, followed by imaging for fluorescence to confirm binding. Proteins were analyzed by electrophoresis for detection and quantification.
ITC assay
Isothermal titration calorimetry (ITC) was performed to analyze interactions of miR5827 with lnc117761 and PKR1 at specific concentrations, assessing binding via dissociation constants.
Ligation of interacting RNA and RT–PCR
A ligation experiment was conducted to investigate binding between lnc117761 and miR5827. The methodology involved crosslinking, circularization, and subsequent amplification to evaluate interactions.
RNA pull-down analysis for binding between lnc117761 and miR5827
Full-length and mutant lnc117761 RNA transcripts were synthesized and used in an RNA pull-down assay to examine the binding affinity for miR5827. The process involved several steps to isolate and quantify the RNA interactions using various techniques.
Dual-luciferase reporter assay
The dual-luciferase reporter assay was set up to assess interactions between RNA sequences and the firefly luciferase gene in rice protoplasts. Measurement of luminescence provided insights into the binding activities in the system.
EMSA
Electrophoretic mobility shift assays (EMSA) were carried out to study interactions between the RNA oligonucleotides and miR5827, using synthesized sequences to visualize binding events on gels.
Uptake of external small RNA by T. aestivum root and coleoptile
Fluorescein-labelled RNA was synthesized and used to analyze uptake in wheat coleoptiles and roots, assessing different treatments and observing RNA distribution under microscope analysis.
Sequence analysis of DNA region encoding RNAs similar to miR5827 from different species
BLAST searches were conducted to find homologous sequences related to miR5827 across various organisms, ensuring a focus on maintaining both the core-binding site and flanking sequences for analysis.
Software and algorithms
Publicly accessible software and algorithms were employed for various analyses including image collection, RNA predictions, and statistical assessments.
Quantification and statistical analysis
Statistical significance was determined using appropriate statistical tests, with careful consideration of sample sizes and data collection techniques to ensure reliability in analyses.
Availability of materials
Constructs, transgenic seeds, and strains generated through this study can be requested, possibly requiring a material transfer agreement.
Reporting summary
Additional details regarding research design can be found in the linked reporting summary.